
Вязко-упругие свойства мыщц разгибателей голеностопного сустава
В 1970 г. Д. Кавагна опубликовал метод определения вязко-упругих свойств икроножной мышцы человека. Испытуемые должны были выполнять небольшой прыжок вверх (20–25 см) с приземлением на вытянутые носки, при этом колени не сгибаются, руки прижаты к выпрямленному телу; испытуемый должен удерживаться на носках в течение 2–3 с.
Прыжки выполняются на динамометрической платформе. Сила, измеряемая на платформе, обусловлена действием сил тяжести и инерции:
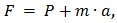
При выполнении прыжков были получены динамограммы с колебанием показателя силы около линии веса. Эти колебания выглядели как затухающие, что и послужило основой для предположения о моделировании тела в виде пружины, массы и вязкого демпфера.
Уравнение движения демпфированного осциллятора имеет вид:
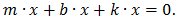
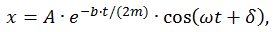
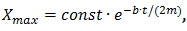 ,
,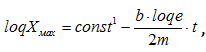
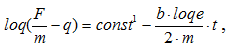
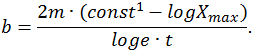
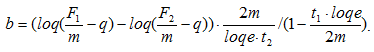
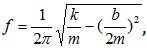
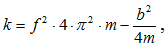
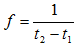 (частота колебаний массы).
(частота колебаний массы).
Пример: пусть t1 = 0,05 c, F1 = 2250 H, t2 = 0,400 c, m = 72 кг, F2 = 1150 H, тогда В = 447 Н/м/с, К = 2,58×104 Н/м.
Эта оценка жесткости в 0,66 раза меньше величины, полученной самим Кавагна (3,8×104 Н/м), но совпадает с оценками, полученными другими исследователями в аналогичном тесте 2,67 ± 0,48×104 Н/м (см. В. М. Зациорский, в соавт., 1981).
Поскольку наши расчеты были выполнены по данным Кавагна, то предполагаем, что им была допущена ошибка в расчетах.
Коэффициент жесткости, полученный таким образом, был пересчитан в коэффициент жесткости для мышц (с учетом плеч действия сил) и составил 13×104 Н/м. Эта оценка оказалась значительно меньше, чем величины, полученные при прямых измерениях на различных мышцах человека. При силе тяги одной мышцы 2250 Н коэффициент жесткости должен быть равен 20×104 или 25×104 Н/м, т. е. в 2,0 раза больше, чем по методике Кавагна. Зависимость «жесткость—сила» имеет линейный вид и может быть представлена следующей формулой К = 125 × F.
Заниженная величина коэффициента жесткости свидетельствует об ошибочности исходных посылок методики Кавагна. Это следует из данных Хилла (1972).
Эффект затухания колебаний активной мышцы нельзя объяснять изменением вязкости мышцы при ее переходе от состояния покоя к активности ... «Все, что известно в настоящее время о мышце, говорит о бессмысленности вязко-упругой теории и модели; лучше всего забыть о них» (Хилл, 1972).
Можно предположить, что по мере растягивания активной мышцы часть мостиков или миофибрилл (при больших напряжениях) разрушается, что и приводит к потере ЗУД.
В чем причина ошибки метода Кавагна?
В рамках программно-целевого подхода ошибочность подхода Кавагна обнаруживается легко. Им был нарушен один из основополагающих принципов научного познания — принцип природной специфичности. Нельзя было по методу аналогии заменять мышцу объектом (моделью), не имеющим сходства по строению и свойствам элементам реальной мышцы.
Попробуем построить более правдоподобную модель объекта — человека, выполняющего описанный выше прыжок.
Человек может быть смоделирован системой, состоящей из трех элементов: первый — туловище и ноги, второй — стопы и третий — мышца.
Свойства элементов модели
Предполагаем, что первый и второй элементы — абсолютно твердые тела, соединенные идеальным цилиндрическим шарниром (без трения). Стопа не имеет массы, а центр масс туловища расположен на высоте Н (100 см) от оси.
Свойства и модель мышцы. Представим мышцу в виде модели, состоящей из сократительного и упругого элементов. Последовательный упругий элемент модели представляет упругие свойства сухожилий и миофибрилл мышечных волокон. Увеличение числа активированных мышечных волокон означает прибавление количества упругих элементов — рост жесткости активной мышцы.
Сократительные компоненты модели функционируют в соответствии с теорией Дещеревского (1968). Взаимодействие между актином и миозином осуществляется посредством мостиков. Мостики находятся в трех состояниях: замкнутые мостики, развивающие тянущую силу; замкнутые мостики, тормозящие скольжение нитей; разомкнутые мостики. Мостики замыкаются независимо друг от друга и тянут нити, вызывая активное сокращение.
Далее те же мостики тормозят движение, так как нити переместились и, наконец, мостики разрываются. Обозначив через n0 полное число активных центров для образования мостиков, через n — число тянущих и m — число тормозящих мостиков, получаем:
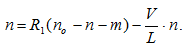
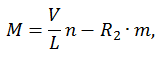
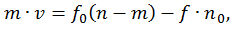
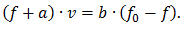
В случае, когда внешняя сила превышает тянущую силу мостиков, сократительный компонент остается в напряженном состоянии. Увеличение внешней силы приводит лишь к удлинению упругих компонентов мышцы, мостики остаются сцепленными, а значит нет затрат энергии (АТФ) на преодоление внешнего сопротивления. При достижении силы, вызывающей резерв сцепленных мостиков, упругие элементы миофибриллы приходят в исходное (ненапряженное) состояние, а энергия упругой деформации рассеивается, превращается в тепло. Поскольку активная мышца не обладает вязкостью, то «сокращение» упругих элементов происходит практически мгновенно (масса мышцы мала) и уже через 1 мс появляются новые мостики. Эти мостики могут преодолеть действие внешней силы, но сначала будут растянуты упругие элементы мышцы в соответствии с действующей силой.
Количество образующихся в мышце мостиков зависит от концентрации ионов в мышечных волокнах. После начала активации мышцы тянущее усилие появляется на мгновение, а только через 25–50 мс достигает максимума, а камбаловидной (медленной мышцы) до 100 мс (Коц, 1982). Развитие активного состояния связывают с увеличением концентрации ионов кальция в саркоплазме мышечных волокон.
Пока активность мышцы растет (увеличивается концентрация кальция) в сократительном компоненте внешнюю нагрузку преодолевают те мостики, которые были образованы до начала растягивания. Одновременно образуются новые мостики, но они не могут взять на себя нагрузку, поскольку им нечего преодолевать. Однако в случае разрыва уже растянутых мостиков появляется нагрузка на образовавшихся мостиках. Заметим, что при разрыве мостиков теряется энергия. Если новых мостиков не образуется, то после разрыва тех, которые преодолевали внешнее сопротивление в мышце, должно снизиться напряжение.
Модель ГСС включает элементы: стопа, туловище, две мышцы. Стопа и туловище являются абсолютно твердыми телами — безинерционными. Они соединены цилиндрическим шарниром без трения. На конце туловища расположена масса.
Мышцы — эквивалентные разгибатели стопы. Первая мышца заменяет (моделирует) трехглавую мышцу голени, вторая мышца заменяет: заднюю, большеберцовую, длинный сгибатель пальцев, длинный сгибатель большого пальца, длинную малоберцовую, короткую малоберцовую.
Первая мышца М-1 крепится на пятке (плечо тяги мышцы L1 = 4 cм в рамках подвижности сустава 60° не меняется).
Вторая мышца имеет другое плечо тяги, L2 она сгибает цилиндрический сустав, поэтому плечо не меняется и составляет 0,2 × L1 = L2 = 0,8 cм.
Предположим, что в ходе движения:
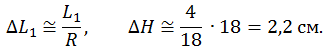
Подъем на 10 см ОЦМ соответствует предельной подвижности голеностопного сустава, дальнейший подъем до плюс 2 см будет связан с деформацией тканей и затем упором костей стопы и голени друг в друга, поэтому 2,2 см + 3 см = предельная величина сокращения М-1, которая согласуется с оценкой растяжения упругой компоненты мышцы 2 см (именно трехглавой, по Александеру, 1977).
Поворот стопы на приводит к уменьшению плеча действия силы реакции опоры, так как вращение идет не вокруг оси, а путем качения — центр давления и ось вращения смещаются вперед одновременно с центром вращения ГСС. Следовательно,
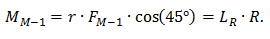
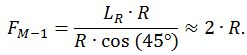
Так как LR = const, то сила тяги мышцы М-2 может быть вычислена
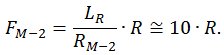
Основные параметры мышц. Мышца М-1 является эквивалентной, заменяет икроножную и камбаловидную. Предположим, что М-1 имеет:
Предположим, что М-2 имеет:
После построения модели необходимо проверить способность ее прогнозировать явления, наблюдаемые в реальном объекте. С этой целью были выполнены четыре эксперимента.
Эксперимент 1
Испытуемый (масса 87 кг) вставал на носки на тензоплатформе четыре раза с разной степенью активации мышц: очень медленно, со средней интенсивностью, быстро, максимально быстро. Регистрировалась реакция опоры. Запись велась на светолучевом осциллографе, скорость протеина ультрафиолетовой бумаги 250 мм/с. Тензоплатформа конструкции ВИСТИ, частота собственных колебаний более 500 ГСС (все последующие эксперименты были выполнены на этой методической базе).
Полученные динамограммы были обработаны с использованием процедуры численного интегрирования. Исходные формулы:
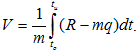
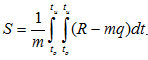
Результаты
По данным эксперимента был рассчитан градиент силы:
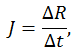
Анализ данных показал, что с увеличением степени активации мышц разгибателей ГСС растет пик силы реакции опоры, линейно увеличивается градиент силы.
Очевидно, в рамках нашей модели мышцы увеличение градиента силы связано с увеличением количества рекрутированных ДЕ (активных мышечных волокон). Градиент J — скорость нарастания напряжения в мышце — зависит от числа активированных МВ, следовательно, удельный градиент должен быть константой.
В формуле:
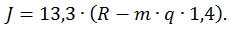
Эта константа должна зависеть от:
Следовательно,
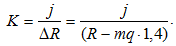
Следующий результат эксперимента. Оказалось, что падение силы реакции опоры наблюдается при одинаковой скорости подъема ОЦМ и сокращения мышцы.
Таблица 1. Путь, скорость и сила реакции опоры при вставании на носки с разной интенсивностью
| № | Интенсивность | Путь ОЦМ, м | Скорость ОЦМ, м/с | Сила, H | Скорость мышц | Мощь, Вт | |
|---|---|---|---|---|---|---|---|
| М-1 | М-2 | ||||||
| 1 | Медленно | 0,049 | 0,46 | 1150 | 0,23 | 0,046 | 529 |
| 2 | Средне | 0,033 | 0,48 | 1280 | 0,24 | 0,048 | 614 |
| 3 | Быстро | 0,025 | 0,44 | 1408 | 0,22 | 0,044 | 619 |
| 4 | Максимально | 0,018 | 0,44 | 1568 | 0,22 | 0,044 | 689 |
Эта скорость для ОЦМ составила 0,44 м/с, для М-1 — 0,22 м/с, для М-2 — 0,044 м/с.
Угловая скорость разгибания стопы в момент пика силы достигла 250 град/с при выполнении упражнения с максимальной интенсивностью, в этот момент мощность разгибания достигла 689 Вт.
При вставании на носки испытуемый поднялся на 10 см (максимальная активность мышц), следовательно, выполнил работу:
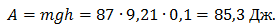
Эксперимент 2
Испытуемый массой в 87 кг выпрыгнул вверх и приземлился на носки напряженных ног.
Результаты
Испытуемый выпрыгнул на 0,159 м. К моменту касания опоры имел скорость ОЦМ 1,77 м/с.
Энергия упругой деформации накапливалась в течение 100 мс. За это время ОЦМ опускался на 11 см, что соответствует удлинению М-1 на 2,2 см. На динамограмме отмечено наложенное колебание силы с частотой 40–50 Гц. При удлинении М-1 с 2,0 см до 2,2 см произошло падение силы реакции опоры, рассеивание энергии. Подъем ОЦМ составил 5,6 см.
За время амортизации испытуемый выполнил следующую работу:
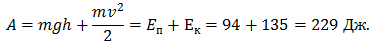
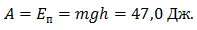
Если предположить, что подъем ОЦМ был выполнен за счет ЗУДМ, то можно рассчитать коэффициент жесткости. Cледовательно, работа могла быть выполнена за счет ПОУК мышц разгибателей голени:
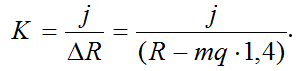
Эксперимент 3
Испытуемый (m = 87 кг) выпрыгивал и приземлялся на носок одной выпрямленной ноги.Результаты
Скорость в момент начала амортизации составила 1,23 м/с, высота падения ОЦМ 9,6 см. Амортизация длилась 140 мс. За это время ОЦМ опустился на 13,7 см. Следовательно, была выполнена работа A = 87 × 9,81 × 0,233 = 199 Дж.За время отталкивания была выполнена работа:
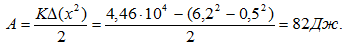
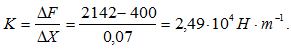
Эксперимент 4
Прыжки вверх на носках без сгибания ног. Испытуемый m=87 кг. Скорость ОЦМ при приземлении составила 1,6 м/с, при вылете 1,56 м/с. За время взаимодействия с опорой была выполнена работа в фазе амортизации или в фазе отталкивания: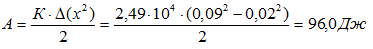
Рассчитаем коэффициент жесткости:
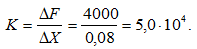
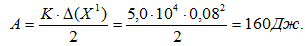
Цель этой статьи — изучить проявление вязких и упругих свойств мышц разгибателей голени. Ответ на вопрос о наличии вязких свойств у мышц был дан Хиллом, ответ, как известно, был отрицателен: нет вязкости, во всяком случае той, которая получается по оценке методики Кавагны или Хилла в 20-х годах. Что же касается упругости, то в трех экспериментах 2, 3, 4, где мышцы функционировали в уступающем режиме, а затем в преодолевающем, были получены неоднозначные результаты.
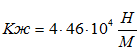
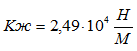
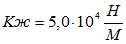
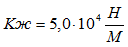
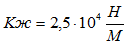
Для случая, когда активная только эквивалентная мышцы М-1 коэффициент жесткости составит:
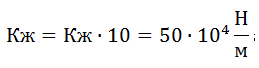 ;
;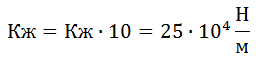 .
.Коэффициенты жесткости, полученные в наших экспериментах, хорошо согласуются с данными литературы (см. В. М. Зациорский с сотр., 1981). На этом можно было бы остановиться, однако обнаруженное нами явление рассеивания энергии в тестах с удержанием позы и отсутствием рассеивания при выполнении прыжков на носках требует объяснения.
Для объяснения этого явления воспользуемся описанной выше моделью голеностопного сустава. Адекватность этой модели был подтверждена результатами первого эксперимента. Например, независимо от степени активации (количества рекрутированных двигательных единиц) максимальная сила и последующее снижение величины опорной реакции наблюдается при одной и той же скорости. Это вызвано тем, что все рекрутированные ДЕ функционируют с максимальной интенсивностью, активация МВ идет одинаково, а увеличение внутреннего трения (числа тормозящих мостиков) зависит только от скорости, видимо, существует индивидуальный оптимум, при котором наблюдается максимум проявляемой силы. Для М-1 этот оптимум соответствует 0,22 м/с, для М-2 этот оптимум должен быть выше, так как мышцы, входящие в этот «эквивалент», более быстрые.
Поэтому при выполнении максимальных прыжков вверх или в беге сократительный компонент М-2 должен выполнять работу, а в М-1 положительную работу может совершить только упругий компонент.
При выполнении прыжков на носках испытуемый в каждом прыжке выполнял отрицательную и положительную работу. В том прыжке, который был зарегистрирован, она составила по 181 Дж. Исходя из данных изменения высоты подъема ОЦМ на опоре (0,08 м) и коэффициента жесткости мышц разгибателей (50×104 Н/м) была вычислена работа, связанная с накоплением и отдачей энергии упругой деформации мышц. Она составила 160 Дж.
Это означает, что основная доля работы приходится на ПОУК. В фазе амортизации была положена энергия в ПОУК 160 Дж, а 21 Дж рассеялся (КПД = 88,4 %). В фазе отталкивания 160 Дж было возвращено ПОУК, а 21 Дж был компенсирован М-2, ее сократительным компонентом. Напомним, что в момент ухода с опоры скорость ОЦМ составила 1,48 м/с, на этой скорости сократительный компонент М-1 работы выполнить не может (468 градусов/с), а сократительный компонент М-2 будет иметь скорость укорочения 0,148 м/с, что значительно меньше найденной в первом эксперименте, «оптимальной» скорости — 0,22 м/с.
Если предположить, что в четвертом эксперименте наши рассуждения правильные, то результаты второго и третьего экспериментов доказывают некорректность наших рассуждений и соответственно неадекватность модели.
Расхождение в экспериментальных данных можно объяснить лишь введением в модель дополнительного элемента — мышцы антагониста (м-3). Если мы введем предположение, что для выполнения задания по удержанию заданной позы (приземлиться и остаться на вытянутых носках стоп) необходимо одновременно активировать как мышцы агонисты (М-1, М-2), так и антагонисты (М-3), то результаты 2-го и 3-го экспериментов становятся понятны.
Физиологический поперечник мышц группы М-3 составляет:
Следовательно, М-3 может проявить изометрическую силу 78,9 × 40 = 3156 Н. Для двух ног 6300 Н.
Эта сила может создавать момент в голеностопном суставе (плечо тяги 0,02 см) 3156 × 0,06 = 189 Н × м.
Следoвательно, если будут одновременно активные все три эквивалентные мышцы, то в уступающем режиме в фазе амортизации доля силы М-3 будет неощутима. Поэтому наши оценки коэффициентов жесткости мышц, вычисленные по силам, возникшим при максимальном растяжении ПОУК, оказались одинаковыми. Иная ситуация возникает при подъеме ОЦМ (фаза отталкивания). В этом случае М-3 работает в уступающем режиме. Выполним работу, необходимую для растяжения М-3:
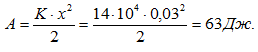
Коэффициент жесткости М-3 был оценен следующим образом: сила изометрической тяги М-1 должна быть равна 22399, при этом Кж = 50 × 104, тогда для М-3 = 6300 Н.
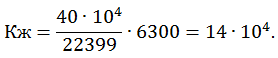
Таким образом, получаем следующий баланс в выполненной работе:
Из представленных расчетов можно видеть, что поведение модели адекватно реальному объекту в случае приближения ее устройства к реальной действительности (с более точным учетом природной специфичности).
Далее для достижения цели, удержания заданной позы целесообразно активировать мышцы агонисты, так и антагонисты. В этом случае система становится более жесткой, амплитуда колебаний быстрее уменьшается.
Результаты, полученные в нашем исследовании, противоречат выводам, сделанным в работе Schenau c cоавт. В их работе было показано, что сократительный компонент мышц разгибателей ГСС не может выполнить работу 191 Дж и создать максимальную мощность 4000 Вт. По оценкам этих авторов, не может выполнить такой работы и ПОУЭ этих мышц, поскольку максимум, что он может дать 50–100 Дж. В связи с этим было введено предположение, что при выполнении волейбольного прыжка голеностопный сустав выполняет работу 190 Дж и достигает мощности 4000 Вт, благодаря передаче мощности от коленного сустава к ГСС через двухсуставную икроножную мышцу.
Видимо, не следует отрицать возможность передачи энергии по двухсуставным мышцам, однако нельзя согласиться с заниженными оценками накопления ЭУД и ПОУК мышц разгибателей голени.
Результаты нашего исследования убедительно показали, что мышцы разгибатели ГСС могут накапливать в ПОУК 160–200 Дж и выполнять преодолевающее движение с максимальной мощностью 4000–5000 Вт.